

سلام به دوستان کنکوری 1401 😊
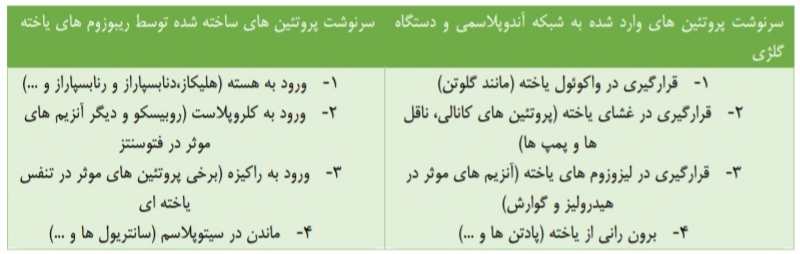
امیدوارم همگی حالتون خوب باشه.
درسنامه و نکات کامل فصل دوم زیست دوازدهم ⬅ جریان اطلاعات در یاخته
منطبق با آزمون ۲۴ دی
نویسنده: آقای محمدمهدی مقصودی، رتبه 268منطقه 2 کنکور تجربی99 و دانشجوی دندانپزشکی دانشگاه اصفهان
درصد زیست کنکور:79%
باماهمراه باشید😄
قسمت دوم
برای دیدن قسمت اول:
صفر تا صد جریان اطلاعات در یاخته_زیست دوازدهم_محمد مهدی مقصودی
میریم برای گفتار دوم و طرح کلی ای از ساخته شدن پلی پپتید!
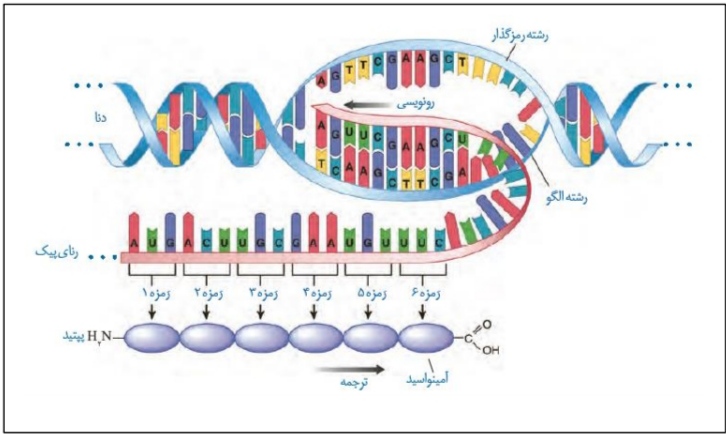
*شکل بالا نشان دهنده رونویسی و سپس ترجمه می باشد.
*به ساخته شدن رشته پلی پپتیدی از روی اطلاعات وراثتی رنای پیک، ترجمه می گوییم.
*یکی از دو رشته دنای مربوط به یک ژن، رشته الگو و رشته دیگر، رمزگذار نامیده می شود.
*همانطور که قبلا هم گفتیم، رونویسی از روی رشته الگو انجام می شود.
*طی فرآیند ترجمه، رنای پیک ترجمه می شود و رشته پلی پپتیدی تولید می شود.
*بر روی رنای پیک توالی های سه نوکلئوتیدی وجود دارد که تعیین کننده آمینواسید های موجود در رشته پلی پپتیدی هستند. به این توالی های سه نوکلئوتیدی رمزه (کدون) می گوییم.
*دقت کنید در یاخته ها، 64 نوع رمزه وجود دارد. اما قرار نیست همه این ها آمینواسید خاصی را رمز کنند. سه نوع رمزه UAA/UGA/UAG هیچ نوع آمینواسیدی را رمز نمی کنند. این رمزه ها، رمزه پایان نامیده می شوند.
*رمزه اول AUG می باشد که رمزه آغاز نیز نامیده می شود. این رمزه مربوط به آمینواسید متیونین می باشد.
*ریبوزوم ها با استفاده از رمزه های سه حرفی رنای پیک، به ترتیب آمینواسید های مختلف را با پیوند پپتیدی به یکدیگر متصل می کنند.
*با توجه به اینکه رمزه آغاز در همه پروتئین های طبیعی AUG می باشد، می توان گفت که اولین آمینواسید در همه رشته های پلی پپتیدی متیونین می باشد.
*دقت کنید که سر آمینی پلی پپتید به آمینواسید میتونین (آغازکننده رشته) و سر کربوکسیل پلی پپتید به آخرین آمینواسید نزدیک تر است.
*قبل از رمزه آغاز و بعد از رمزه پایان نیز توالی هایی وجود دارد، اما دقت کنید که این توالی ها ترجمه نمی شوند!
*از آنجایی که آمینواسید ها در جانوران مختلف یکسان هستند، جانوران دارای رمزه های مشابه در ماده وراثتی خود هستند.
*با توجه به محصور نبودن دنای پروکاریوت ها در هسته، فرایند رونویسی و ترجمه می تواند همزمان انجام شود.
*دقت کنید که گفتیم رنای تازه رونویسی شده، ممکن است نابالغ باشد و نیاز به تغییراتی داشته باشد. این مراحل و تغییرات در شکل نشان داده نشده است.
در شکل بعدی، می خواهیم بریم سراغ ساختار رنای ناقل!
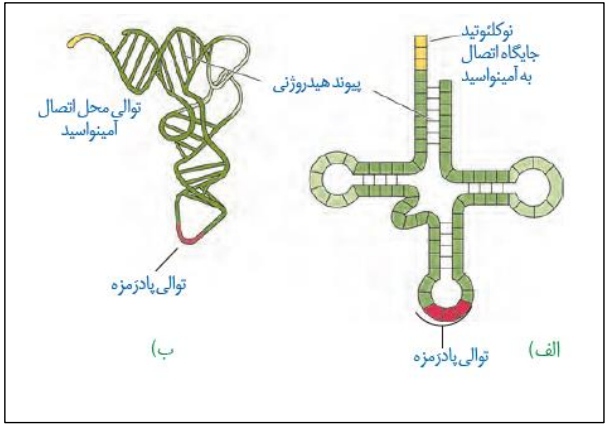
*در گفتار قبل راجع به برخی تغییراتی که روی رنای پیک صورت می گرفت، صحبت کردیم. در این شکل به بررسی تغییراتی که روی رنای ناقل انجام می شود، می پردازیم.
*شکل الف) این شکل تاخوردگی اولیه رنای ناقل را نشان می دهد. در واقع تشکیل پیوند هیدروژنی بین بخش های خاصی از رنای ناقل، سبب تاخوردگی اولیه می شود.
*شکل ب) این شکل نشان دهنده ساختار نهایی و سه بعدی رنای ناقل است. با تاخوردگی های بیشتر ساختار الف، این ساختار ایجاد می شود. در واقع این ساختار است که فعالیت می کند، به آمینواسید متصل می شود و در پروتئین سازی شرکت می کند.
*ساختار نهایی سه بعدی رنای ناقل، L شکل می باشد.
*ساختار رنای ناقل بالغ، شامل چهار بازو و سه حلقه می باشد. از میان حلقه ها، دو حلقه آن جانبی و یک حلقه مرکزی است. توالی آنتی کدونی بر روی حلقه میانی قرار گرفته است.
*دقت کنید بین نوکلئوتیدهای تشکیل دهنده حلقه ها، پیوند هیدروژنی تشکیل نمی شود.
*در بازوی منتهی شده به حلقه مرکزی، یک برآمدگی مشاهده می شود.
*در مقابل توالی آنتی کدونی، محلی برای اتصال آمینواسید وجود دارد. همانطور که در شکل می بینید، نوکلئوتید های این بخش فاقد مکمل می باشند و توانایی تشکیل پیوند هیدروژنی را ندارند. در واقع سه نوکلئوتید جایگاه اتصال آمینواسید هستند که البته همه آمینواسید ها به قند ریبوز آخرین نوکلئوتید (از این توالی) متصل می شوند.
*دقت کنید ساختار رنا های ناقل به جز در توالی آنتی کدونی، مشابه می باشد. بنابراین تعداد پیوند هیدروژنی تشکیل شده درون خود رنای های ناقل یکسان می باشد.
*اتصال آمینواسید به آخرین نوکلئوتید توالی مربوطه به کمک تشکیل پیوند کوالانسی خواهد بود!
*حلقه های جانبی در ساختار اولیه روبروی هم ولی در ساختار رنای بالغ در کنار هم هستند!
*رنای ناقل در یوکاریوت ها به وسیله رنابسپاراز 3 و در پروکاریوت ها توسط رنابسپاراز پروکاریوتی ساخته می شود.
*همواره اولین رنای ناقلی که درون ریبوزوم قرار می گیرد دارای توالی آنتی کدونی UAC است و حامل میتوین می باشد.
*ترکیب با 5 یازدهم: دقت کنید در یاخته هایی مانند لنفوسیت های پادتن ساز و ... که در شرایطی پروتئین سازی افزایش می یابد، میزان رناهای ناقل موجود در یاخته زیاد می شود.
گفتیم که رنای ناقل در واقع حمل کننده آمینواسید است! در این شکل می خواهیم ببینیم چجوری؟
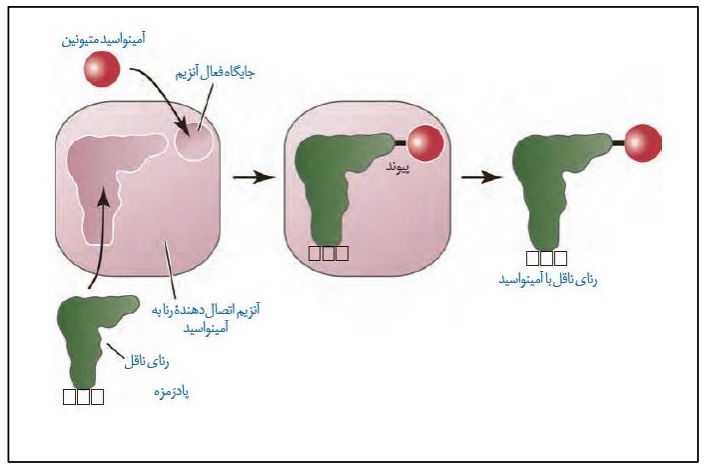
*در یاخته ها آنزیمی وجود دارد که سبب اتصال آمینواسید به رنای ناقل می شود.
*آنزیم مربوطه دارای دو جایگاه مشابه با ساختار آمینواسید و رنای ناقل را درون خود دارد. در واقع هر
دو مولکول مربوطه کاملا روی آنزیم جای می گیرند.
*آنزیم مربوطه با شناسایی توالی آنتی کدونی رنای ناقل، آمینواسید مربوطه را پیدا می کند و به رنای ناقل متصل می سازد.
*این آنزیم در حین فعالیت خود، نیازمند انرژی است که از ATP تامین می شود. با استفاده از این انرژی پیوند اشتراکی بین آمینواسید و رنای ناقل برقرار می سازد.
*با توجه به اینکه بیشتر آمینواسید ها دارای چندین کدون هستند، می توان گفت یک آمینواسید می تواند توسط چند نوع رنای ناقل حمل شود ولی هر رنای ناقل تنها یک نوع آمینواسید را منتقل می کند.
*ترکیب با 5 دوازدهم:آنزیم متصل کننده رنای ناقل به آمینواسید همانند آنزیم سازنده ATP از کراتین فسفات دارای دو پیش ماده می باشد.
گفتیم که پروتئین سازی توسط ریبوزوم ها صورت میگیره! در این شکل می خواهیم با ساختارش آشنا بشیم...
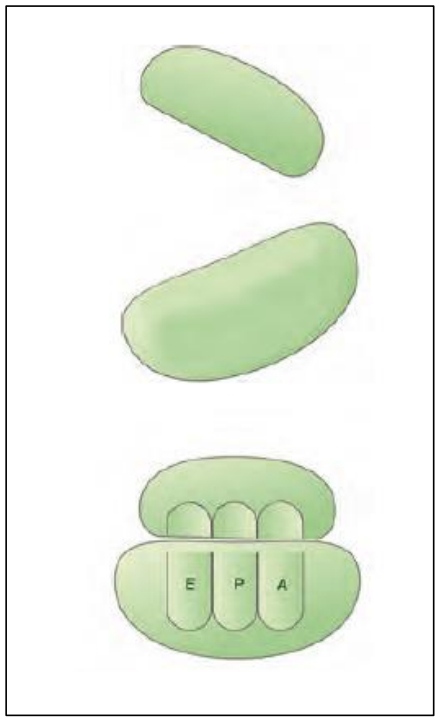
*ریبوزوم ها که در ساخت پلی پپتیدی ها نقش دارند، از دو زیرواحد کوچک و بزرگ ایجاد شده اند.
*ریبوزوم ها از ساختار های پروتئینی و نوکلئیک اسیدی(rRNA) تشکیل شده اند. در واقع می توان گفت حداکثر 24 نوع مونومر در ساختار آن نقش دارند. (20 نوع آمینواسید و 4 نوع نوکلئوتید)
*در ساختار ریبوزوم کامل سه جایگاه E/P/A یافت می شود. بخش عمده این جایگاه ها در زیرواحد بزرگ قرار گرفته است.
*در ساختار ریبوزوم ها، رنای رناتنی وجود دارد که فعالیت آنزیمی دارد. این آنزیم سبب برقراری پیوند پپتیدی بین آمینواسیدها می شود.
*رنای رناتنی قرارگرفته در ساختار ریبوزوم، دارای شکل تاخورده و ساختار سه بعدی می باشد.
*رنای رناتنی توسط رنابسپاراز 1 در یوکاریوت ها و در پروکاریوت ها توسط رنابسپاراز پروکاریوتی ساخته می شود.
*دقت کنید که ریبوزوم ها تنها سبب ایجاد ساختار اول پروتئین ها می شوند. بقیه ساختار بعدا با روشی که در فصل قبل بیان کردیم، ایجاد می شوند.
*در مراحل ترجمه نیز می گوییم که زیرواحد کوچک و بزرگ ریبوزوم ها تنها در هنگام ترجمه در کنار یکدیگر قرار می گیرند. در سایر زمان ها، جدا از هم دیگر یافت می شوند.
*ترکیب با 5 دوازدهم: دقت کنید که درون میتوکندری ها و کلروپلاست ها نیز ریبوزوم وجود دارد. البته دقت کنید که این ریبوزوم ها نسبت به ریبوزوم های موجود در سیتوپلاسم دارای اندازه کوچکتر و ساختار ابتدایی تر می باشند.
این همه گفتیم و گفتیم تا رسیدیم به ترجمه! توی سه شکل بعدی می خواهیم این فرایند رو در سه مرحله بررسی کنیم:
![]()
*در مرحله آغاز، ابتدا زیرواحد کوچک ریبوزوم به رنای پیک متصل می شود. (این کار به وسیله توالی هدایت کننده ای که قبل از AUG (رمزه آغاز) وجود دارد، صورت می گیرد.)
*در گام بعدی، رنای ناقل دارای توالی آنتی کدونی UAC (رنای ناقل حمل کننده متیونین) در سمت دیگر رنای پیک به آن متصل می شود.
*در نهایت زیرواحد بزرگ ریبوزوم به مجموعه مربوطه متصل می شود و ساختار رناتن به صورت کامل ایجاد می شود.
*همانطور که در شکل قبلی هم گفتیم، در ساختار کامل ریبوزوم سه جایگاه E/P/A دیده می شود. در مرحله آغاز تنها زیرواحد P اشغال شده است.
*دقت کنید که در مرحله آغاز، پیوند پپتیدی ای تشکیل نمی شود و ریبوزوم حرکت نمی کند.
*مطابق شکل، لزوما کدون آغاز در ابتدای رنای پیک وجود ندارد و ممکن است توالی هایی قبل از آن نیز یافت شوند.
*دقت کنید زمانی که رنای ناقل آغازگر با کدون آغاز پیوند هیدروژنی برقرار می کند، هنوز ساختار ریبوزوم کامل نشده است.
*مطابق شکل سمت راست، رنای ناقل تنها در مقابل زیرواحد بزرگ دیده می شود.
*تنها پیوندی که در مرحله آغاز تشکیل می شود، پیوند هیدروژنی است! در واقع در این مرحله تشکیل پیوند پپتیدی نداریم.*ممکن است در رنای پیک بیشتر از یک کدون AUG وجود داشته باشد، اگر جاندار یوکاریوت باشد تنها اولین AUG کدون آغازگر نامیده می شود. اما اگر با پروکاریوت ها طرف باشیم، بیش از یکی آن ممکن است کدون آغازگر باشد! این موضوع را در گفتار سوم همین فصل بیشتر متوجه می شویم:)
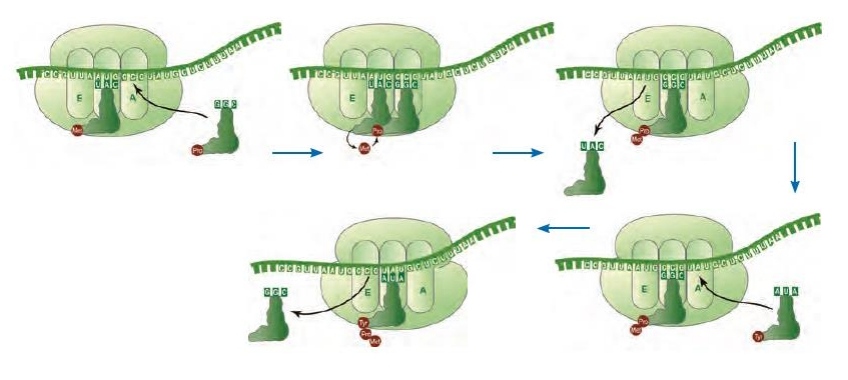
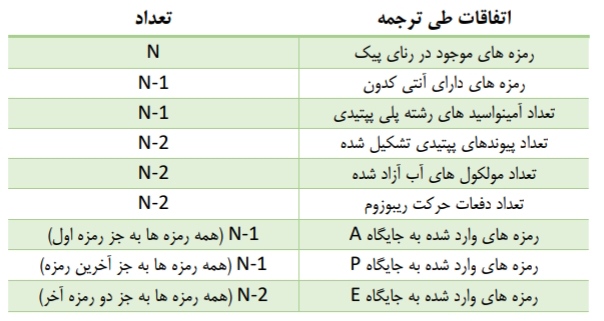
*در مرحله طویل شدن، رناهای ناقل مختلفی وارد جایگاه A می شوند، اما تنها یکی از آنها که توانایی برقرار پیوند هیدروژنی با رمزه جایگاه A را دارد، مستقر می شود.
*در گام بعدی، آمینواسید متصل به رنای ناقل که در جایگاه P قرار گرفته است، از رنای ناقل جدا می شود و با اتصال به آمینواسید متصل به رنای ناقل در جایگاه A، پیوند پپتیدی ایجاد می کند.
*پس از اینکه دو گام بالا انجام شد، ریبوزوم به اندازه یک کدون (سه نوکلئوتید) جلوتر می رود. با اینکار، رنای ناقل موجود در جایگاه A به جایگاه P، رنای ناقل موجود در جایگاه P وارد جایگاه E می شود. با تکرار این گام ها، مرحله طویل شدن انجام می شود.
*در ارتباط با نکته قبل دقت کنید که ابتدا رنای ناقل فاقد آمینواسید از جایگاه E خارج می شود، سپس رنای ناقل حاوی آمینواسید وارد جایگاه A می شود!
*دقت کنید بعد از حرکت اول، در حرکت های بعدی، رنای ناقل موجود بعد از دادن آمینواسید خود و واردشدن به جایگاه E، از طریق این جایگاه، رنای ناقل را ترک می کند.
*نخستین پیوند پپتیدی در ابتدای مرحله طویل شدن انجام می شود.دقت کنیم تنها در این مرحله، تشکیل پیوند پپتیدی را شاهد هستیم.
*در این مرحله، 5 توالی سه نوکلئوتیدی کدون و آنتی کدون به طور همزمان درون جایگاه های ریبوزوم مشاهده می شود.
*برخلاف اولین رنای ناقل که وارد جایگاه P می شود، همه دیگر رناهای ناقل ابتدا به جایگاه A وارد می شوند.
*کدون آغاز و اولین رنای ناقل تنها در جایگاه های E و P حضور دارند، اما سایر رناهای ناقل درون هر سه جایگاه می توانند مشاهده شوند.
*آخرین حرکت ریبوزوم روی رنای ناقل، پس از تشکیل آخرین پیوند پپتیدی در جایگاه A رخ می دهد.
*خروج رشته پلی پپتیدی در این مرحله همواره از جایگاه P صورت می گیرد. در واقع از این جایگاه خارج می شود و به جایگاه A وارد می شود.*ترکیب با 1 دوازدهم: دو گروه آمین و کربوکسیل آمینواسید های مختلف می توانند به همدیگر نزدیک شوند و با خروج یک مولکول آب، تشکیل پیوند پپتیدی دهند. پیوند پپتیدی در واقع نوعی پیوند اشتراکی محسوب می شود.
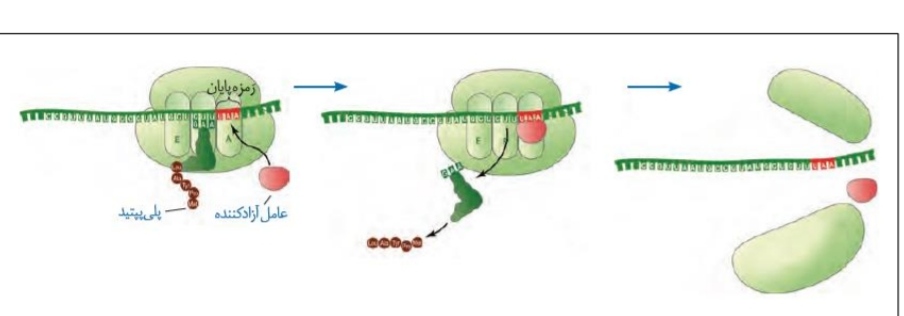
*با ورود یکی از کدون های UGA/UAG/UAA به جایگاه A ریبوزوم، عمل ترجمه پایان می پذیرد.
*هنگامی که یکی از کدون های بالا در جایگاه A قرار می گیرد، پروتئین هایی با عنوان کلی عامل آزادکننده وارد این جایگاه می شوند و سبب جداشدن رشته پلی پپتیدی ساخته شده از جایگاه P می شود. در گام بعدی، خود رنای ناقل نیز از همین جایگاه جدا می شود.
*پس از اینکه رنای ناقل و رشته پلی پپتیدی جدا شدند، دو زیرواحد ریبوزوم از یکدیگر جدا می شوند.
*عوامل آزادکننده تنها در زیرواحد بزرگ ریبوزوم دیده می شوند.
*عوامل آزادکننده، پروتئینی هستند و توانایی ایجاد رابطه مکملی و پیوند هیدروژنی با کدون های پایان را ندارند.
*واضح است که در مرحله پایان هیچ رنای ناقلی وارد جایگاه A نمی شود و آخرین رنای ناقل از جایگاه P خارج می شود.
*کدون و آنتی کدون آغازگر تنها در جایگاه های E و P مشاهده می شوند، در حالی که دیگر رمزه ها به جز رمزه پایان و آنتی کدون پایانی، قطعا وارد هر سه جایگاه می شوند.رمزه پایان تنها در جایگاه A دیده می شود و آنتی کدون پایانی (آنتی کدون مربوط به کدون ماقبل آخر) تنها در جایگاه های P و A مشاهده می شود.
در آخر جمع بندی داریم از بحث ترجمه و نکاتی که گفتیم:
در شکل بعدی به این موضوع می پردازیم که خب، رشته پلی پپیتیدی ساخته شد! حالا کجا میره و چه اتفاقاتی روش رخ میده!؟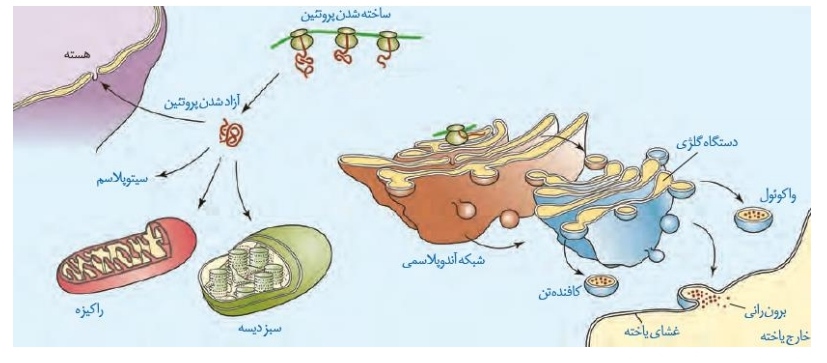 *تعدادی از ریبوزوم های یاخته پس از ساخت بخش کوچکی از رشته پلی پپتیدی به شبکه آندوپلاسمی متصل می شوند و بقیه رشته را در این حالت می سازند.
*تعدادی از ریبوزوم های یاخته پس از ساخت بخش کوچکی از رشته پلی پپتیدی به شبکه آندوپلاسمی متصل می شوند و بقیه رشته را در این حالت می سازند.
*رشته هایی که ریبوزوم های گفته شده می سازند، پس از ساخت ابتدا به شبکه آندوپلاسمی و سپس به دستگاه گلژی وارد می شوند.
*پروتئین هایی که به دستگاه گلژی وارد می شوند، بسته بندی می شوند و سرنوشت های مختلفی دارند.
*دسته ای از این پروتئین ها به درون واکوئول ها، دسته ای به درون کافنده تن ها و دسته دیگر با برون رانی از یاخته خارج می شوند.
*ریبوزوم های دیگر موجود در سیتوپلاسم پس از ساخت رشته، آن را رها می کنند.
*این دسته از پروتئین ها می توانند به درون هسته، راکیزه ها و یا کلروپلاست ها بروند. دسته دیگر این پروتئین ها به صورت آزاد درون سیتوپلاسم قرار می گیرد.
*توالی های آمینواسیدی خاصی در پروتئین ها وجود دارد که تعیین می کند پروتئین به کجا باید برود که نمونه هایی از آن را گفتیم!
*پروتئین هایی که به سمت اندامک های دو غشایی (شامل هسته، راکیزه و کلروپلاست) می روند، درون دسته گلژی بسته بندی نمی شوند.
*پروتئین های غشایی از مسیر شبکه آندوپلاسمی و دستگاه گلژی عبور می کنند.
*ریبوزوم های متصل به شبکه آندوپلاسمی از طریق زیرواحد بزرگ خود به آن متصل شده اند.
*بخشی از شبکه آندوپلاسمی یاخته در اطراف هسته و بخشی از آن در نزدیک غشای یاخته دیده می شود.
*بخش های کیسه ای شکل شبکه آندوپلاسمی برخلاف دستگاه گلژی به یکدیگر متصل هستند.
*آنزیم هایی مانند هلیکاز، دنابسپاراز و رنابسپاراز که در این دو فصل راجع بهشون صحبت کردیم، توسط ریبوزوم های آزاد درون سیتوپلاسم ساخته می شوند.
*ترکیب با 5 یازدهم: پلاسموسیت ها دارای قدرت زیاد برای تولید پادتن (نوعی پروتئین ترشحی) هستند، مطابق شکل آنها در کتاب یازدهم این یاخته ها دارای شبکه آندوپلاسمی و دستگاه گلژی وسیعی هستند. پادتن ها از طریق این مسیر تولید می شوند.
*ترکیب با 6 یازدهم: ریزکیسه های سازنده تیغه میانی در گیاهان توسط دستگاه گلژی بسته بندی می شوند. این ریزکیسه ها طی آنافاز تا پایان تقسیم سیتوپلاسم به یکدیگر متصل می شوند.با ادغام این ریزکیسه ها، دیواره یاخته ای شکل می گیرد.
*ترکیب با 7 یازدهم: ریزکیسه های سازنده جدارلقاحی تخمک توسط گلژی بسته بندی می شوند و در هنگام لقاح محتویات خود را با برون رانی بیرون می ریزند.
*ترکیب با 5 و 6 دوازدهم: دقت کنید که درون راکیزه و سبزدیسه ها نیز ریبوزوم ها وجود دارد. بنابرین می توان گفت پروتئین های موجود در این اندامک ها دو مبدا دارند، یا درون خود اندامک مربوطه ساخته شده اند و یا توسطریبوزوم های آزاد درون سیتوپلاسم تولید شده اند.
در آخرین شکل گفتار می خواهیم به بررسی سرعت و مقدار ترجمه بپردازیم!
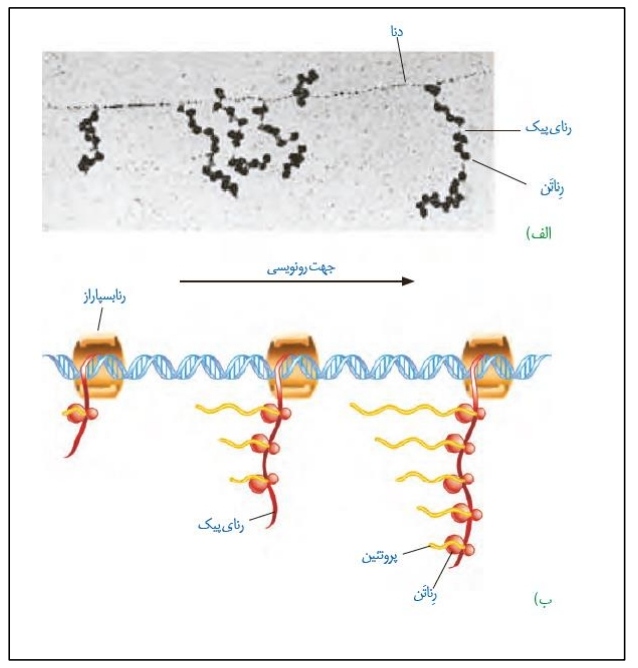
*سرعت و مقدار پروتئین سازی برحسب نیاز یاخته در زمان های مختلف متفاوت می باشد.
*شکل الف نشان دهنده یک مولکول دنا می باشد که همزمان چندین رنابسپاراز در حال رونویسی و ایجاد رنای پیک هستند. روی این رنای پیک به طور همزمان، چندین ریبوزوم در حال ترجمه دیده می شود.
*ساختار ایجاد شده در شکل بالا، در پروکاریوت ها دیده می شود. به دلیل اینکه طول رنای پیک در این یاخته ها کم است، ساخت پروتئین ها توسط مجموعه ای از ریبوزوم ها به طور همزمان و پشت سر هم صورت می گیرد.
*به دلیل تجمع ریبوزوم ها، ساختاری شبیه تسبیح ایجاد می شود که در آن رنا مانند نخ تسبیح و دانه های تسبیح همان ریبوزوم های متصل به رنای پیک می باشند.
*تجمع رناتن ها (پلی ریبوزومی) در یاخته های یوکاریوتی نیز دیده می شود.
*رنابسپاراز هایی که زودتر به دنا متصل شده اند، رونویسی بیشتری انجام داده و در واقع به توالی پایان نزدیک تر می باشند.
*جهت رونویسی از سمت رناهایی با طول کمتر به سمت رناهایی با طول بیشتر می باشد.
*مشخص است که ریبوزوم هایی که زودتر شروع به فعالیت کرده اند، پلی پپتید بلندتری ساخته اند. این ریبوزوم ها مطابق شکل به رنابسپاراز در حال رونویسی از دنای یاخته نزدیک تر هستند.
*هر چه در جهت رونویسی حرکت کنیم، تعداد زنجیرهای پلی پپتیدی و ریبوزوم های در حال ترجمه بیشتری را مشاهده می کنیم.
در اولین شکل گفتار سه به بررسی تنظیم منفی رونویسی در باکتری ایکلای می پردازیم! قند ترجیحی این باکتری گلوکز هست ولی در شرایطی که گلوکز نباشد، باکتری می تونه از لاکتوز استفاده کنه! این قند با گلوکز فرق میکنه و خب طبعن آنزیم های لازم برای متابولیسمش هم متفاوته! اما حواستون باشه که اگر در محیط باکتری لاکتوز وجود نداشته باشه یا مقدارش کم باشه، آنزیم های تجزیه کننده اون هم متوقف میشن یا کاهش پیدا میکنن!
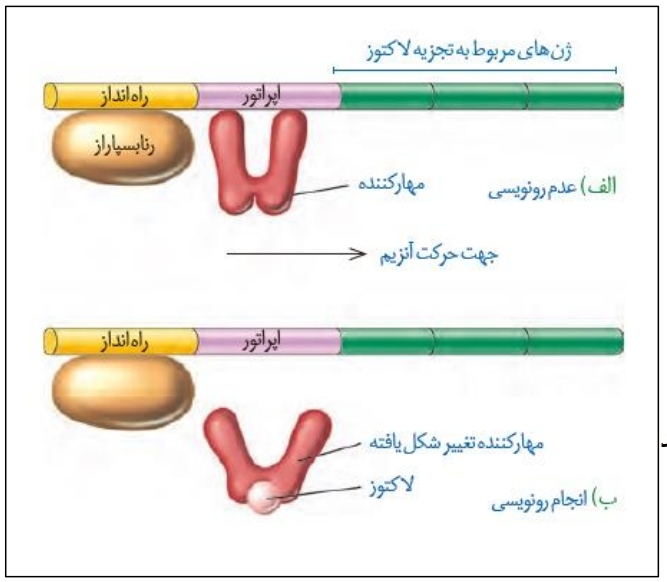
*به طور کلی اگر مانعی بر سر فعالیت آنزیم رنابسپاراز وجود داشته باشد، این نوع تنظیم را تنظیم منفی رونویسی می گوییم.
*در تنظیم منفی رونویسی، در حضور گلوکز پروتئینی به نام مهار کننده به توالی بین ژنی خاصی به نام اپراتور متصل می شود.
*وقتی که مهارکننده به اپراتور متصل می شود، رنابسپارازی که به توالی راه انداز متصل شده است، توانایی حرکت را ندارد و در نتیجه رونویسی از ژن های مربوط به تجزیه لاکتوز رخ نخواهد داد.
*اگر در محیط باکتری، لاکتوز برخلاف گلوکز حضور داشته باشد، این قند وارد باکتری می شود و به پروتئین مهارکننده متصل می شود.
*با اتصال لاکتوز به پروتئین مهارکننده، این پروتئین تغییر شکل می دهد و بدین سبب از توالی اپراتور جدا می شود. مطابق شکل، تغییر شکل پروتئین مهارکننده به این صورت است که دو بازوی آن از یکدیگر دور می شوند!
*با جداشدن پروتئین مهارکننده از اپراتور، مانعی در برابر رنابسپاراز دیگر نیست و این آنزیم از روی اپراتور رد می شود و بعد از آن رونویسی از اولین ژن مربوط به تجزیه لاکتوز را انجام می دهد.
*برای تجزیه لاکتوز نیاز به محصولات سه ژن داریم. این سه ژن در کنار یکدیگر قرار گرفته اند و راه انداز مشترکی دارند!
*ژن اول دارای نوکلئوتید جایگاه آغاز رونویسی و ژن آخر هم دارای توالی پایان رونویسی می باشد. پس دقت کنید لزوما هر ژنی دارای نوکلئوتید آغاز رونویسی و توالی پایان رونویسی را ندارد.
*دقت کنید همیشه و همواره رنابسپاراز به راه انداز متصل است. در واقع این اتصال فارغ از وجود لاکتوز یا عدم وجود آن می باشد.
*با توجه به اینکه اگر لاکتوز وجود داشته باشد، پروتئین مهارکننده به آن متصل می شود می توان نتیجه گرفت که تمایل اتصال پروتئین مهارکننده به لاکتوز بیشتر از اتصال به دنا می باشد.
*رنای ساخته شده مربوط به آنزیم های تجزیه کننده لاکتوز، دارای سه کدون آغاز و سه کدون پایان می باشد. این رنای پیک پس از ترجمه، سه رشته پلی پپتیدی متفاوت را ایجاد می کند.
*دقت کنید هر ژن مربوطه به ساخت یک آنزیم است، در واقع محصولات هر یک از ژن ها با دیگری متفاوت است.
*جایگاه آغاز رونویسی در تنظیم منفی رونویسی به توالی اپراتور متصل است، نه راه انداز!
*همیشه اینگونه نیست که ژن به توالی راه انداز خود متصل باشد، گاهی مانند آنچه که دیدیم توالی های دیگری مانند اپراتور می تواند بین آنها قرار گیرد.
*مطابق شکل اندازه توالی اپراتور و راه انداز از هریک از ژن های موثر در تجزیه لاکتوز بزرگتر هستند.
*ژن مربوط به ساخت پروتئین مهارکننده در جای دیگری از دنای باکتری است و رونویسی از آن و تولید این پروتئین ربطی به وجود گلوکز یا لاکتوز ندارد.
*ترکیب با 1 دهم: لاکتوز دی ساکاریدی است که از گلوکز و گالاکتوز ایجاد شده است.
در شکل قبلی راجع به تنظیم منفی رونویسی صحبت کردیم، در این قسمت می خواهیم به سراغ تنظیم مثبت بریم!
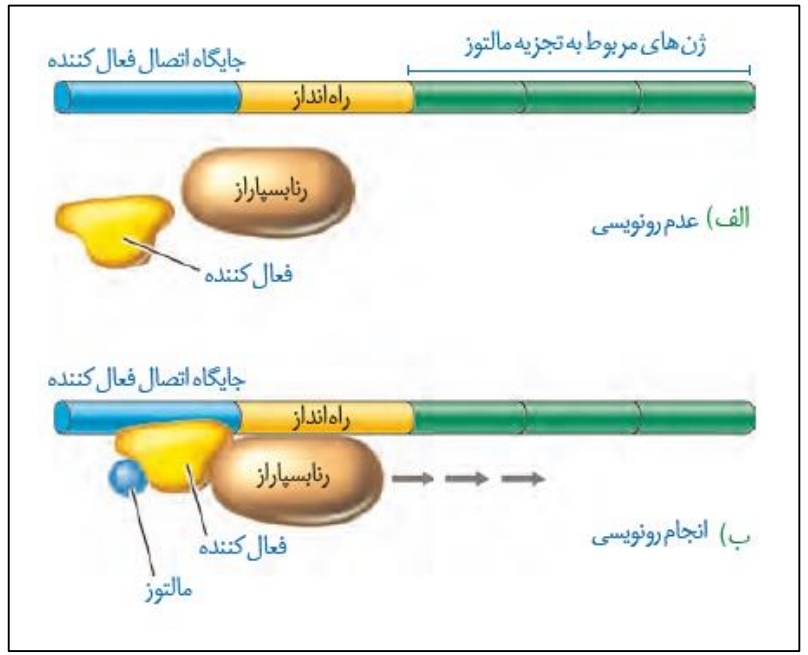
*اگر پروتئین های خاصی در یاخته به رنابسپاراز کمک کنند تا بتواند به راه انداز متصل شود، این نوع رونویسی از تنظیم مثبت رونویسی می گوییم.
*در شرایطی که مالتوز در یاخته وجود داشته باشد، این مولکول قندی به بخش خاصی از پروتئین
فعال کننده متصل می شود و باعث اتصال این پروتئین به توالی بین ژنی تنظیمی ای به نام جایگاه اتصال فعال کننده می شود.
*پروتئین فعال کننده متصل به جایگاه اتصال فعال کننده سبب می شود که رنابسپاراز به توالی راه انداز متصل شود و بدین ترتیب رونویسی صورت می گیرد.
*مطابق شکل، توالی جایگاه اتصال فعال کننده قبل از راه انداز قرار دارد، به همین دلیل رنابسپاراز از روی آن عبور نمی کند.
*با توجه به نکته قبل، جایگاه اتصال فعال کننده مورد رونویسی واقع نمی شود!
*همانند لاکتوز، سه ژن برای آنزیم های تجزیه کننده مالتوز داریم. ژن اول دارای نوکلئوتید آغاز رونویسی و ژن آخر دارای توالی پایان رونویسی می باشد.
*دقت کنید مالتوز همانند لاکتوز مستقیما به دنا متصل نمی شود. لاکتوز به پروتئین مهار کننده و مالتوز به پروتئین فعال کننده متصل می شود.
*پروتئین فعال کننده برخلاف پروتئین مهارکننده هنگام اتصال به دی ساکارید تغییرشکل نمی دهد!
*توالی اپراتور در مجاورت جایگاه آغاز رونویسی قرار داشت ولی جایگاه اتصال فعال کننده در مجاورت نوکلئوتید آغاز رونویسی قرار ندارد.
*پروتئین فعال کننده توانایی اتصال به سه نوع موکول مختلف (قند مالتوز، پروتئین آنزیمی رنابسپاراز و دنا) را دارد ولی پروتئین مهارکننده تنها به دنا و قند لاکتوز می تواند متصل شود.
*همانند تنظیم منفی، رنای پیک مربوط به سه ژن ساخته می شود که با ترجمه، سبب ساخت سه نوع آنزیم برای تجزیه مالتوز می شود.
*در تنظیم منفی رونویسی اتصال رنابسپاراز به راه انداز بدون نیاز به پروتئین خاصی صورت می گیرد ولی در تنظیم مثبت، رنا بسپاراز به فعالیت پروتئین فعال کننده لازم دارد.
*مشابه مهار کننده، رونویسی از ژن های مربوط به ساخت فعال کننده ربطی به حضور یا عدم حضور مولکول های قندی ندارد.
*ترکیب با 1 دهم: مالتوز مولکول قندی دی ساکاریدی است که از دو واحد گلوکز تشکیل شده است.
تا اینجای کار شما با نمونه هایی از تنظیم رونویسی در پروکاریوت ها آشنا شدین، در این قسمت میخواهیم بریم سراغ تنظیم رونویسی در یوکاریوت ها!
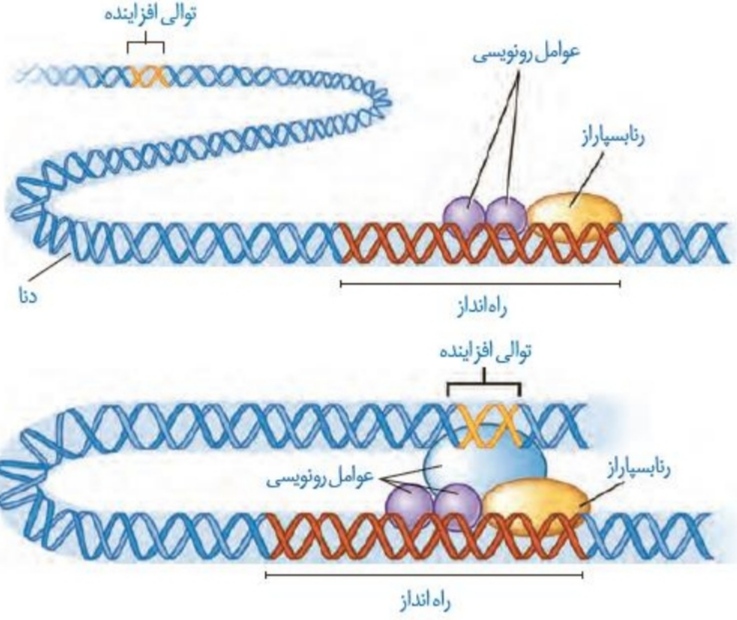
*در یوکاریوت ها، رنابسپاراز نمی تواند به خودی خود توالی راه انداز را شناسایی کند و نیاز به پروتئین هایی با عنوان کلی عوامل رونویسی هستند.
*عوامل رونویسی به نواحی خاصی از راه انداز (در اواسط آن) متصل می شوند و در گام بعدی به رنابسپاراز کمک می کند تا به راه انداز متصل شود.
*رنابسپاراز به سمتی از راه انداز متصل می شود که به ژن مربوطه نزدیک تر است.
*میل اتصال عوامل رونویسی به راه انداز در اثر عواملی تغییر می کند، به همین دلیل مقدار و سرعت رونویسی در شرایط مختلف متفاوت می باشد.
*در یوکاریوت ها برخلاف پروکاریوت ها، بین راه انداز یک ژن و خود ژن فاصله ای وجود ندارد.
*به جز عوامل رونویسی متصل شونده به راه انداز، عوامل رونویسی دیگری وجود دارند که به توالی تنظیمی خاصی به نام افزاینده متصل می شوند.
*در اثر اتصال عوامل رونویسی به توالی افزاینده، مولکول دنا خم می شود. با خم شدن مولکول دنا، عوامل رونویسی متصل به راه انداز و توالی افزاینده در کنار یکدیگر قرار می گیرند که این موضوع سبب افزایش سرعت و مقدار رونویسی می شود.
*مطابق شکل، عوامل رونویسی متصل به توالی افزاینده نسبت به عوامل رونویسی متصل به راه انداز و حتی خود رنابسپاراز بزرگ تر می باشند.
*دقت کنید برخی عوامل رونویسی به راه انداز متصل نمی شوند، مانند عوامل رونویسی متصل به توالی افزاینده!
*رنابسپاراز و عوامل رونویسی متصل به افزاینده تنها با بعضی عوامل رونویسی متصل به راه انداز اتصال برقرار می کنند.
*وقتی برخی عوامل رونویسی به توالی افزاینده متصل می شوند، خمیدگی در دنای یاخته رخ می دهد.
*دقت کنید ایجاد خمیدگی در دنا مربوط به یوکاریوت هاست نه پروکاریوت ها!
*توالی افزاینده نسبت به راه انداز می تواند اندازه کوچک تری داشته باشد.(از نوکلئوتیدهای کمتری تشکیل شده است.)
*توالی افزاینده می تواند در فاصله دوری نسبت به ژن قرار داشته باشد و لزوما مانند راه انداز در کنار آن قرار نگرفته است.
برای رزرو مشاوره و رفع اشکال درسی و شرکت در کلاس های کنکوری با مشاوران سایت برترها بر روی لینک زیر کلیک کنید یا با شماره تماس 0216463 داخلی1174(حیدریان) تماس بگیرید.سایت برترها